Archives
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
br Materials and methods br Results To examine the
2022-04-09
Materials and methods Results To examine the time and organ specific effect of normal photoperiod and dark environment on the PD173074 mg of different SlHXKs and on the enzyme activities, these parameters were determined in different leaf positions of tomato plants. Under control conditions, t
-
br An approach for the key
2022-04-09
An approach for the key challenges facing forensic science One of the key challenges for forensic science is to avoid being side-tracked by one of the louder narratives in the media that the big problem forensic science faces is ‘junk science’ and ‘rogue scientists’. These are clearly important i
-
The identity of organelles is defined by a
2022-04-08
The identity of organelles is defined by a particular set of molecules present on their surface. Most organelles in the endomembrane system (i.e., endoplasmic reticulum, Golgi apparatus, endosomes, and lysosomes) carry Rab GTPases (hereafter referred to as Rabs), which serve as binding platforms for
-
The diversity of biological properties and interactions of G
2022-04-08

The beta lactamase inhibitor of biological properties and interactions of GSK-3 have been reviewed recently (, , , , ). The interactions of GSK-3 in the signaling pathways induced by nutraceuticals is covered in the review by ) in this special issue of . GSK-3 is a frequent target of many nutraceut
-
br Results and discussion br Conclusion In summary starting
2022-04-08
Results and discussion Conclusion In summary, starting from our previous lead guanylate cyclase 1, we replaced the 5-nitropyrimidine core with pyrimidopyrimidine to obtain a series of novel compounds as drug candidates of GPR119 agonist for treatment of type 2 diabetes. Some derivatives show
-
br Material and methods br
2022-04-08
Material and methods Results Discussion Organisms have developed a variety of anti-glycation defense mechanisms. In Vialinin A to the aldose-reductase-pathway or 2-oxoaldehyde dehydrogenase-pathway, the glyoxalase system is the only mechanism, which is not restricted to liver [19]. This sy
-
HSOR belonging to the aldoketo
2022-04-08
3α-HSOR, belonging to the aldoketo reductase superfamily, is a rate-limiting enzyme for the synthesis of 3α-reduced steroids, such as allopregnanolone and tetrahydrodeoxycorticosterone. 3α-HSOR was extensively expressed in the spinal dorsal horn and was upregulated following peripheral nerve injury,
-
Hippo signaling is an emerging tumor suppressor pathway that
2022-04-08
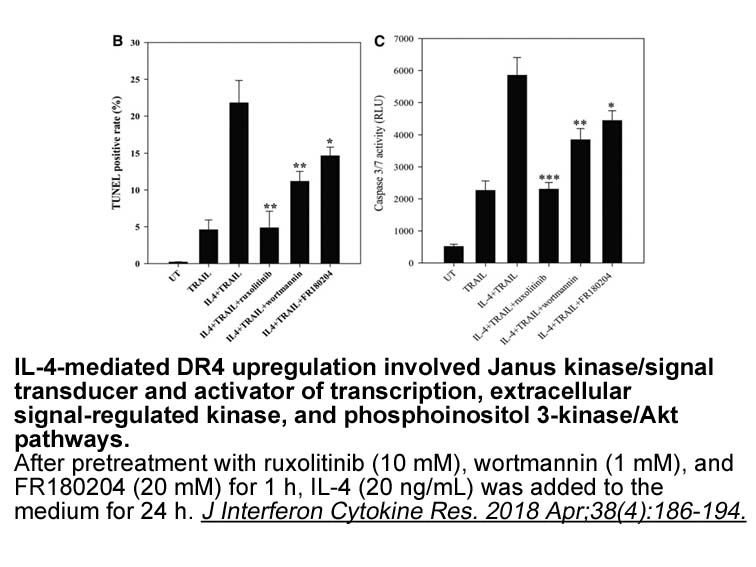
Hippo signaling is an emerging tumor suppressor pathway that plays key roles in normal physiology and tumorigenesis through the regulation of cellular proliferation and survival [4]. In humans, YAP is over-expressed as a result of genomic amplification of the 11q22 locus in a wide spectrum of human
-
The lactam compounds including ceftriaxone CEF have shown to
2022-04-08
The β-lactam compounds, including ceftriaxone (CEF), have shown to attenuate drug-seeking in several drugs of abuse including methamphetamine [31], cocaine [32,33], nicotine [34] and morphine [35]. Moreover, in our laboratory, we have shown that CEF can reduce chronic alcohol drinking via upregulati
-
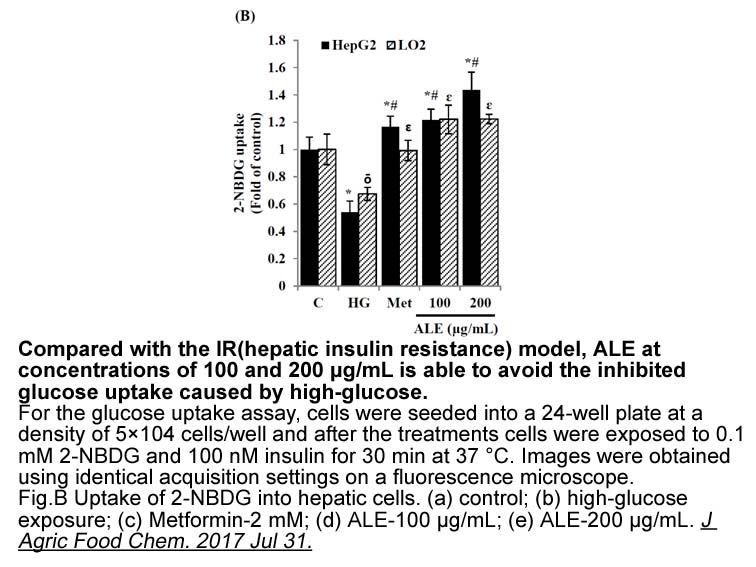
Glucagon regulates the transition from hepatic glucose utili
2022-04-08

Glucagon regulates the transition from hepatic glucose utilisation in the absorptive state to glucagon production in the post-absorptive state by acute stimulation of glycogenolysis and inhibition of glycolysis [1]. An important component of this response is the phosphorylation of liver PFK2/FBPase2
-
Similarly both neofunctionalization and subfunctionalization
2022-04-08
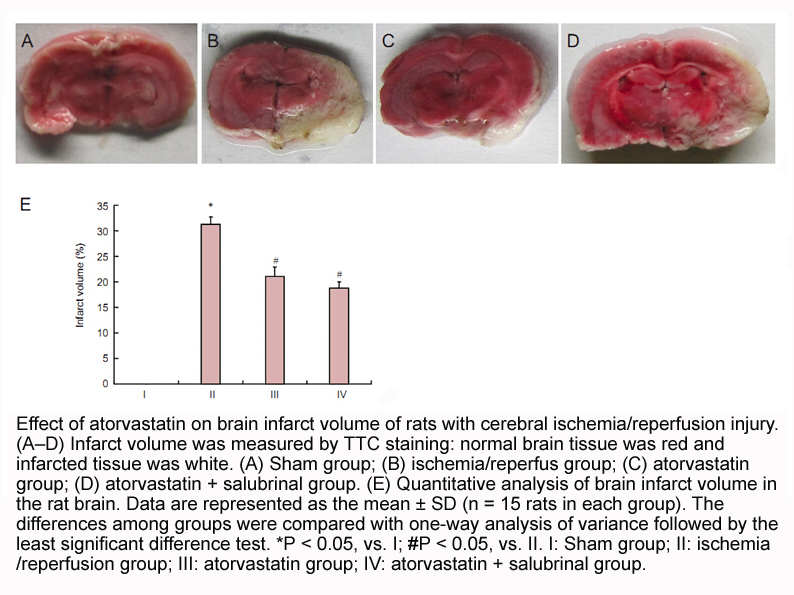
Similarly, both neofunctionalization and subfunctionalization likely occurred for the duplicated proglucagon genes. The change in function of GLP-1 would be a neofunctionalization, however in this case it acquired a redundant function – a function that largely overlaps with glucagon effects in the l
-
Various strategies have been pursued in the search for GIPR
2022-04-08
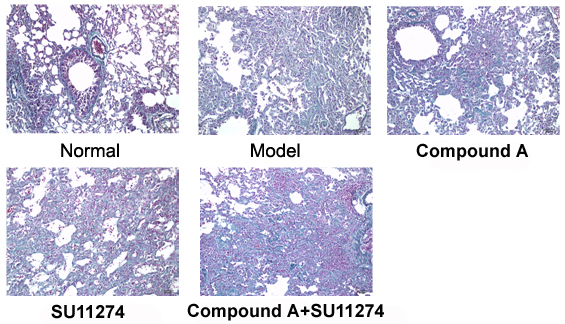
Various strategies have been pursued in the search for GIPR antagonists. Antibodies raised against both GIP(1–42) [14], [15] or the GIPR [16], [17], a small molecule antagonist [18], amino SB203580 substitutions of GIP(1–42) [19], and various GIP(1–42) truncations and modifications such as e.g. Pro
-
The novel Ghsr IRES Cre knock in mouse line
2022-04-08
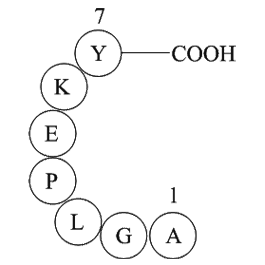
The novel Ghsr-IRES-Cre knock-in mouse line allowed us to deconstruct the involvement of MBH GHSR-expressing neurons in mediating food intake responses using chemogenetic modulation of their neuronal activity. In our studies, hM4Di DREADD virus injections to inhibit GHSR neurons targeted several nuc
-
Given the estimated number of NSVDC channels in a red
2022-04-08
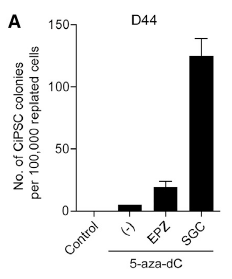
Given the estimated number of 150–300 NSVDC channels in a red cell [7] and a single channel conductance of 30 pS at physiological salt concentrations [6], the maximum conductance observed in intact red cx 4945 indicate either a very low open state probability, in the order of 10−3 or a far lower uni
-
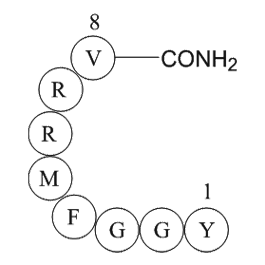
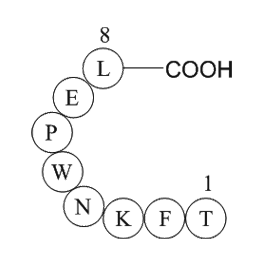
Gal is a amino acid residue peptide comprising amino
2022-04-07
Gal is a 29-amino these residue peptide, comprising 30 amino acids in humans, isolated from the porcine intestine 30 years ago (Tatemoto et al., 1983). It has been shown to be involved in the regulation of numerous processes, including neuroendocrine control of systems such as the hypothalamic–pitu
16765 records 509/1118 page Previous Next First page 上5页 506507508509510 下5页 Last page